ISSN Number
ISSN 2771-019X-

-

Impact Factor
1.2*
ISSN Number
ISSN 2771-019X
Impact Factor
1.2*1Department of Geriatric Medicine, Affiliated Hospital of Guilin Medical University, China.
2Department of Respiratory and Critical Care Medicine, Second Affiliated Hospital of Guilin Medical University, China.
#These authors have been equally contributed to this article.
Department of Respiratory and Critical Care Medicine, Second Affiliated Hospital of Guilin Medical University, China.
Tel: +86-07732824373;
Email: 247546160@qq.com
Received : Jan 18, 2025,
Accepted : Feb 20, 2025
Published : Feb 27, 2025,
Archived : www.jclinmedcasereports.com
Background and aims: The effects of brain death and cardiac death on lung tissue were different. In the pathological process of brain death and heart death, the activation and regulation of complement system show obvious differences. Thus, our study aims to explore the difference of complement-related genes between the two.
Methods: Using data from the Gene Expression Omnibus (GEO), we explored the expression profiles of Complement-Related Genes (CRGs) in lung tissues from Donate after Brain Death (DBD) and Donate after Cardiac-Dead (DCD) with the help of the R data package. Our study involved analyzing protein interactions, investigating gene correlations, and conducting functional enrichment. Key genes were identified using a range of machine learning techniques.
Results: Analysis of GEO datasets, which included 24 DBD and 11 DCD samples, identified two critical complement-related genes (CRGs): UNC119, FBLN2, and BCHE. The combination of these genes resulted in an Area Under the Curve (AUC) of 0.993, effectively distinguishing between lung tissues from DBD and DCD. Additionally, nomogram, decision curve, and calibration curve analyses confirmed their diagnostic efficacy.
Conclusion: We created a prognostic model for lung tissues from DBD and DCD donors using two pivotal Complement-Related Genes (CRGs), which showed impressive predictive power. These insights deepen our comprehension of the complement system’s role in DBD and DCD lung tissues.
Keywords: Donation after brain death; Donation after cardiac death; Complement; Machine learning.
Copy right Statement: Content published in the journal follows Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0). © Jiangfa L (2025).
Journal: The Journal of Clinical and Medical Images, Case Reports (JCMICR) is a fantastic resource for keeping up with the latest clinical advancements and for publishing case reports and clinical images related to a variety of medical illnesses.
The effects of brain death and cardiac death on lung tissue were different. Death is usually accompanied by neurogenic pulmonary edema and systemic inflammatory response, which is caused by the excessive release of norepinephrine from sympathetic nerve stimulation, resulting in increased permeability of pulmonary microvessels, resulting in acute pulmonary edema [1,2]. At the same time, people with brain death are often supported by mechanical ventilation, which can maintain oxygen supply while avoiding organ ischemia [3]. However, after cardiac death, rapid circulatory arrest leads to ischemia-reperfusion injury and rapid hypoxia of lung tissue, which leads to a range of cellular damage and metabolic dysfunction, increasing the risk of pulmonary edema and inflammatory responses. In this case, donated lungs may not be as stable in texture as those obtained at brain death, and recovery after transplantation is difficult due to unmanageable ischemic time [4-6]. Therefore, understanding the different effects of the two on lung tissue is critical to optimizing strategies for the acquisition and preservation of transplanted organs.
The complement system is an important part of the innate immune response and plays multiple roles in inflammatory responses and tissue damage [7]. It consists of a series of plasma proteins that are activated in response to pathogen invasion or tissue damage, initiating expansion responses via the classical, bypass, and lectin pathways. Complement activation leads to increased cell lysis and phagocytosis, in which C3 and C5 complement components play a key role in regulating the inflammatory response by attracting neutrophils and other immune cells to the site of injury or infection. In addition, the complement system can directly cause pathogen destruction through the formation of Membrane Attack Complexes (MAC). However, overactivation or improper regulation of the complement system can lead to excessive inflammatory responses and tissue damage, exacerbating pathological damage and triggering diseases such as autoimmune disease and transplant rejection [8]. Therefore, the complement system plays a double-edged role in maintaining the balance between immune defense and damage repair.
In the pathological process of brain death and heart death, the activation and regulation of complement system show obvious differences. Activation of the complement system in brain-dead states is often strongly associated with Systemic Inflammatory Response Syndrome (SIRS), which may be due to significant changes in norepinephrine/epinephrine levels triggered by brain death. At this time, complement activation may exacerbate the inflammatory response, resulting in increased damage to lung and other tissues [9]. However, because the heart still keeps beating, complement-mediated damage is relatively better controlled. In contrast, after cardiac death, lung tissue is at risk of ischemia-reperfusion injury due to the rapid cessation of blood circulation, and the complement system is dramatically activated during reperfusion, resulting in a massive release of inflammatory cytokines. In this case, the complement system may further exacerbate reperfusion-related tissue damage and cell necrosis [10]. Therefore, the activation pathways of the complement system and its contribution to tissue damage in the two states of death show distinct patterns, and this difference needs to be deeply understood and managed during organ protection and transplantation.
Patients and datasets
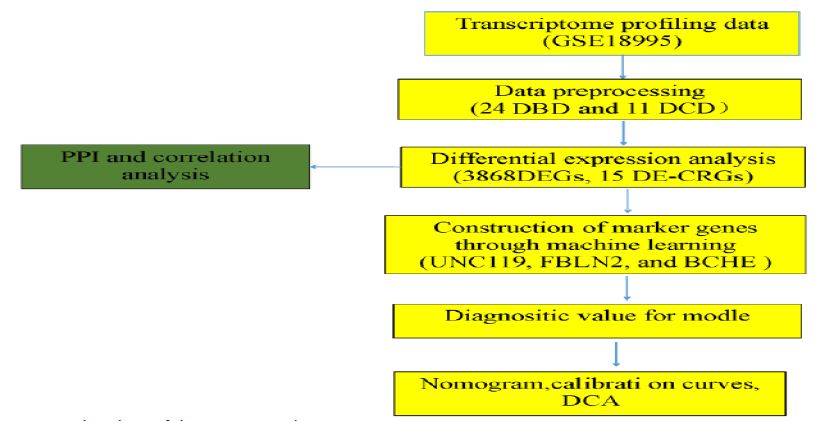
The transcriptomic analysis of lung specimens from DBD and DCD donors utilized the GSE18995 dataset, obtained from the GEO database, which included 24 DBD and 11 DCD samples. The flowchart illustrating this study is presented in (Figure 1).
Expression of DEGs and CRGs between DBD and DCD
We used the R package “limma” to perform a differential expression analysis on the processed data from the GEO database, identifying Differentially Expressed Genes (DEGs) between DBD and DCD. The results were visualized with volcano and heatmap plots using the “ggplot2” and “heatmap” R packages, with the criterion for selection being an adjusted P<0.05. The intersection of DEGs (|log CF| >1 and adjusted P<0.05) related to CRGs was identified using the “VennDiagram” R package and defined as DE-CRGs for further analysis. A boxplot for the DE-RGs was created using the “ggpubr” R package. A Venn diagram was used to pinpoint CRGs within DRGs between DBD and DCD samples. Subsequently, multivariate analysis was conducted to identify key DE-CRGs.
Correlation analysis and protein‒protein interaction (PPI) network construction
The R package “heatmap” was used to generate heatmaps for 15 DE-CRGs. PPI networks for these DE-CRGs were constructed using the STRING database (https://string-db.org/). Furthermore, we employed the “ggplot2” package (version 3.3.6) in R to conduct pairwise correlation analysis of the dataset variables, visualizing the results in a heatmap.
Gene Ontology (GO) and KEGG pathway enrichment analysis
To analyze the biological function of genes, we employed the “clusterProfiler” package in R, which facilitated the enrichment analysis of Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways [11-13]. The GO annotation encompassed three domains: Biological Processes (BP), Cellular Components (CC), and Molecular Functions (MF).
Machine learning methods to identify key genes
The “glmnet” package in R was used to conduct Least Absolute Shrinkage and Selection Operator (LASSO) regression on the chosen linear model, effectively reducing data dimensionality while retaining essential variables [14]. Furthermore, we integrated Random Forest (RF) algorithms for optimal gene selection. RF, a regression tree technique, leverages bootstrap aggregation and randomization of predictors to achieve high prediction accuracy, implemented via the “randomForest” package in R [17]. The genes identified by these three machine learning methods were intersected to establish the final key genes.
The Establishment of the the DE-CRGs diagnostic Model
In developing the DE-CRGs diagnostic model, we utilized the GSE18995 dataset. Initially, we constructed a nomogram model using the “rms” package in R to predict the likelihood of DBD occurrence. Following this, we employed the “pROC” package [18] to assess the Area Under the Curve (AUC), along with the specificity and sensitivity of the diagnostic value for the marker genes through time-dependent ROC analysis. Each central gene was assigned a score, which was then aggregated to produce a total score.
Statistical analyses
Continuous variables are reported as mean ± standard deviation. The student’s t-test was used for comparisons between two groups, while the Wilcoxon rank-sum test was applied to non-normally distributed variables. A p-value of less than 0.05 was considered statistically significant. The symbols *, **, and *** indicate p-values of less than 0.05, 0.01, and 0.001, respectively. All statistical analyses were conducted using R software (version 4.2.1).
Identification of DE-CRGs associated with DBD
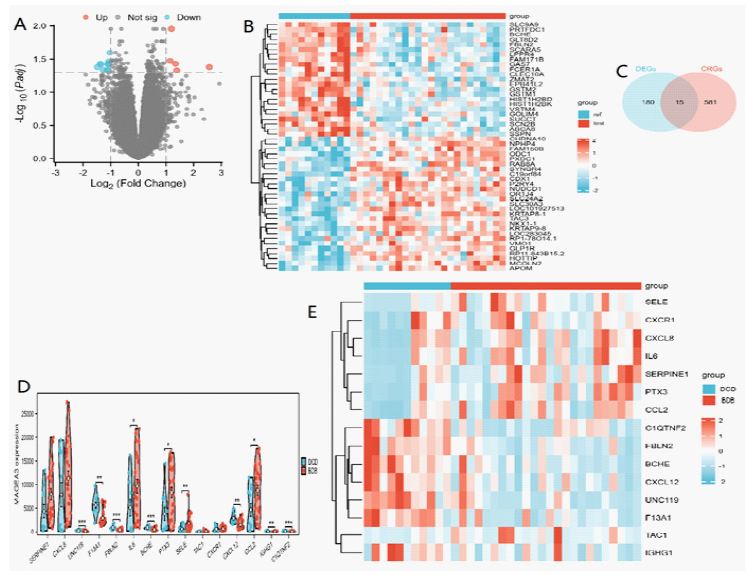
Using the “limma” package, we identified 3,868 DEGs (adj. p<0.05) from the GSE18995 dataset, which included 24 DBD and 11 DCD samples. Among these, 86 genes were up-regulated, and 109 were down-regulated. The volcano plot illustrating the differentially expressed genes is presented in (Figure 2A), while (Figure 2B) displays the heatmap of the top 50 differential genes between DBD and DCD. Additionally, 596 CRGs [19] overlapped with the 195 DEGs((|log CF| >1 and adj. p<0.05), revealing 15 DE-CRGs with significant differences between the DBD and DCD (Figure 2C). Eight DE-CRGs (TAC, CXCR, SERPINE, CXCL8, PTX3, CCL2, IL6, and SELE) were high expression and seven DE-CRGs (C1QTNF2, UNC119, F13A1, CXCL12, FBLN2, BCHE, and IGHG1) were low expression in DBD (Figure 2D), and the heatmap of these 15 DE-CRGs is shown in (Figure 2E).
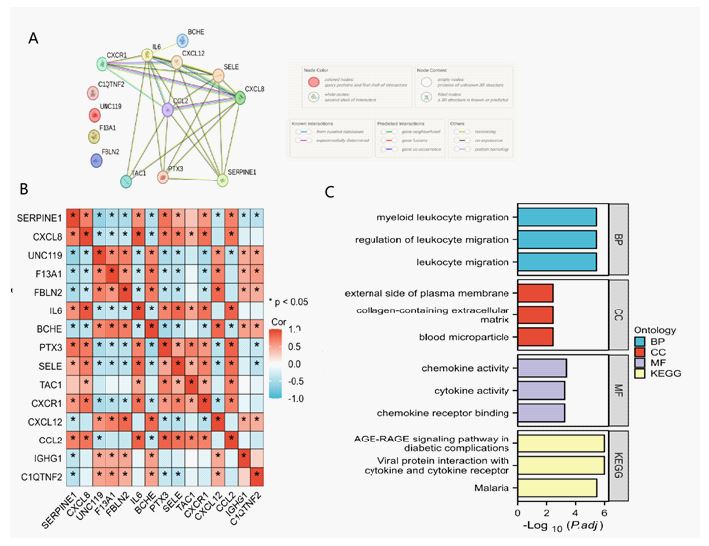
A PPI analysis was conducted using STRING to examine potential interactions among the 15 Differentially Expressed Complement-Related Genes (DE-CRGs), as shown in (Figure 3A). The correlations among these 15 DE-CRGs are presented in Figure 3B. DE-CRGs were found to be related to response to Lipopolysaccharide (LPS), inflammasome complex, cysteine-type endopeptidase activity, and cytokine receptor binding, among other pathways, as revealed by GO enrichment analysis (Figure 3C). KEGG pathway analysis showed involvement in lipid and atherosclerosis and non-alcoholic fatty liver disease (Figure 3C).
Identification of diagnostic marker genes for DBD
Considering the individual complexity and heterogeneity of DBD and DCD patients, candidate CRGs were identified from 15 DE-CRGs using RF and LASSO validated machine learning models to assist in predicting DBD diagnosis (Figures 4A & B). Out of the 15 DE-CRGs analyzed, three were successfully identified (refer to Figure 4C).
Evaluation of the diagnostic marker genes
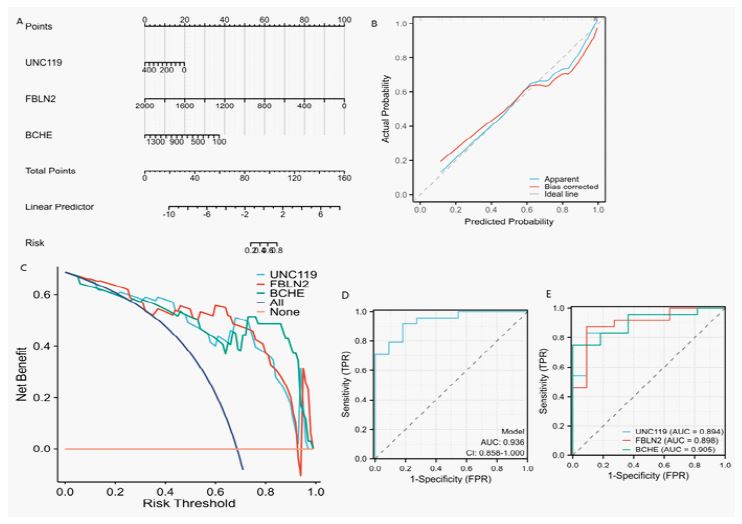
A nomogram model was created to differentiate lung tissue from DBD and DCD, incorporating three central genes: UNC119, FBLN2, and BCHE (Figure 5A). The numerical values of each biomarker in the nomogram were used to distinguish between the two tissue types, and a calibration curve showed a strong correlation between predicted and actual probabilities (Figure 5B). Decision Curve Analysis (DCA) indicated that the net benefit from this model was significantly greater than zero, underscoring its considerable accuracy and utility in clinical decision-making (Figure 5C). ROC curve analysis demonstrated that the combined features of the three key genes performed well in distinguishing between DBD and DCD lung tissues (AUC=0.936, Figure 5D), with individual predictive ROC results for each gene closed to 0.90 (Figure 5E). These results suggest that the model based on these three marker genes has strong predictive efficacy for differentiating between lung tissues from DBD and DCD.

Brain Death (BD) donors are the main organ source for lung transplantation [20]. The death process can induce acute lung injury and aggravate lung ischemia-reperfusion injury [15]. While many immune mechanisms have been shown to stimulate donor organ damage and predispose grafts to poor outcomes, activation of the complement system has been shown to play a central role in BD-related graft damage. In both preclinical and clinical studies of heart and kidney transplants, grafts from BD donors were not only associated with more severe Ischemia-Reperfusion Injury (IRI), but posttransplant pathology was also significantly associated with levels of complement activation [21,22].
Complement is a major component of the immune system [23]. It is heavily involved in the body’s defense function and its own immune regulation, and its overactivation plays a priming and amplifying effect in the initial stage of inflammatory response. The complement system is a particularly important upstream sensor and effector system, and the activation products of complement are important regulators in inflammatory and immune processes. As an important component of the immune system, complement is heavily involved in the body’s defense response and its own immune regulation, and its overactivation plays an initiating and amplifying effect in the initial stage of inflammatory response [23]. Many causes of ALI have been experimentally demonstrated to be involved in the activation of the complement system [24].
In this study, we investigated the potential role of CRGs in lung tissue of DBD and DCD, identified potential key genes. We retrieved lung tissue samples from DBD and DCD patients from the GEO database for statistical analysis to identify DEGs, ultimately identifying 15 DEGs associated with complement pathways. These results imply that CRGs may play a role in the progression of lung injury. Our correlation analysis showed that the identified DE-CRGs were closely interconnected; however, some exhibited no clear correlation at the protein level, emphasizing the heterogeneity in CRG interactions at both gene and protein levels.
The important role of DE-CRGs in response to myeloid leukocyte migration, regulation of leukocyte migration, external side of plasma membrane, collagen-containing extracellular matrix, chemokine activity chemokine receptor binding, viral protein interaction with cytokine and cytokine receptor, AGE-RAGE signaling pathway in diabetic complications, and malaria was revealed by GO and KEGG enrichment analyses.
Analyses using LASSO, and RF of the 15 DE-CRGs identified three key genes—UNC119, FBLN2, and BCHE—that effectively differentiate lung tissue from DBD and DCD, achieving an AUC of 0.936. The nomogram model, supported by calibration curves and Decision Curve Analysis (DCA), demonstrated robust predictive capability and significant clinical relevance.
UNC119 is a conserved intracellular protein that mainly plays a role in the nervous system, especially in neurons [25], and is involved in a variety of biological processes such as cell signal transduction, cytoskeleton regulation and membrane transport. UNC119 binds to some intracellular small molecules (such as small GTPase), regulates their activity, and plays an important role in the development and function of neurons [26]. UNC119 may regulate the function of immune cells by affecting intracellular signal transduction pathway, thereby indirectly affecting the activation and regulation of complement [27]. In immune response, UNC119 may be involved in the interaction between immune cells and complement components, affecting their ability to recognize and clear pathogens. UNC119 may play a regulatory role in the development and function of immune cells, thereby affecting complement-related immune response. FBLN2 (Fibulin-2) is an extracellular matrix protein, belonging to the fibronectin family, which plays an important role in tissue development, repair and maintenance. FBLN2 plays a key role in a variety of biological processes, including cell adhesion, signaling, and the formation of tissue structure [28-30]. FBLN2 may affect the function of complement system by influencing the structure of extracellular matrix and altering the deposition and activation process of complement components. FBLN2 is expressed in certain immune cells, such as macrophages and lymphocytes, and may influence their response to complement activation by regulating the adhesion and migration of these cells. Butyrylcholinesterase (BCHE) is an enzyme in blood plasma that is primarily responsible for the hydrolysis of cholinesterase compounds [31] in the body, BCHE plays an important role in neurotransmission, drug metabolism, detoxification and other processes. Similar to Acetylcholinesterase (AChE), BCHE acts mainly in some tissues close to neurons, including blood and liver, but it has a broader substrate specificity. The main physiological function of BCHE is to hydrolyze Butyrylcholine and other related ester compounds, which play an important role in the regulation of neurotransmission, although its role is relatively limited compared with AChE. The activation of the complement system usually triggers an inflammatory response, and BCHE may play a supporting role in this process, by hydrolyzing and clearing some inflammatory mediators, alleviating the intensity of the inflammatory response. This also hints at its potential role in regulating complement-mediated inflammation [32].
Our study has some limitations. Firstly, we conducted genetic analysis on data sourced from the GEO database, which may introduce certain biases. Secondly, the sample size was relatively small, which could impact the generalizability of the findings. Finally, we did not conduct additional tests to verify the expression of these genes.
We initially identified two significant genes that, in combination, accurately differentiate between lung tissue from DBD and DCD.
Ethics approval and consent to participate: The patient datas used in the article was downloaded from a public database, so the approval of the unit ethics committee and the participant’s signed consent were waived.
Consent for publication: The patient datas used in the article was downloaded from a public database, so the participant’s consent for publication were waived.
Availability of data and materials: The datasets in this study were enrolled from the GEO database (https://www.ncbi.nlm.nih.gov/geo/), with the following data accessions enrolled: GSE18995. The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Competing interests: The authors declare no conflicts of interest in relation to the research, indicating that the study was conducted without any commercial or financial relationships that could be perceived as potential conflicts of interest.
Funding: This study was supported by. Self-funded Project of Guangxi Zhuang Autonomous Region Health Commission (Z20200261).
Author’s contributions: LPL, and JFL conducted the formal analysis and initial draft of the manuscript, with project administration being overseen by LPL, and JFL performed software analysis. Data curation was handled by JFL, and LPL, while the execution of experiments was carried out by LPL, and JFL all contributed to the writing of the article. All authors participated in the editing process and approved the manuscript for submission.
Acknowledgements: We are grateful to the researchers who provided the datasets (GSE18995). We thank the website (https://www.xiantaozi.com) for some data analysis on this website.
